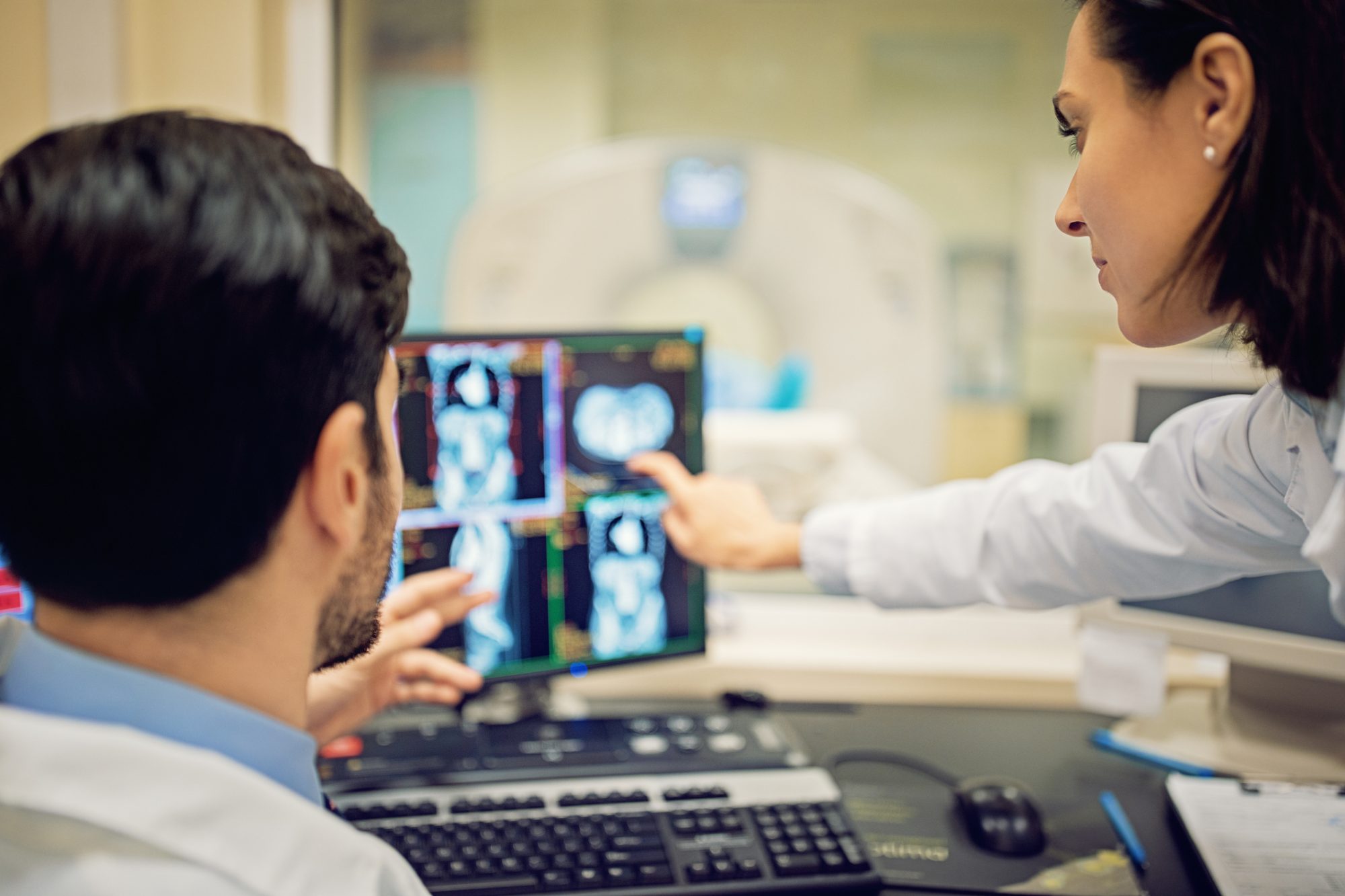
Interdisciplinary approaches in developmental biology have revealed how cells build the embryonic bones of the skull vault. Spatial and temporal dynamics are coordinated by cells as they build the extracellular environment, Jacqueline Tabler explains
Cell collectives build form
An organ’s function is fundamentally tied to the way its tissue is built. Every feature of an organ, including shape, durability, or its capacity for repair, is determined by collectives of cells and their actions in space and time. Exploring how cell collectives build organs, therefore, offers new insights into organ biology, evolution, and disease.
The skeleton exemplifies the inextricable link between cells and the form that determines function in vertebrates throughout their lives. (1,2) Osteoblasts (bone-forming cells) secrete a mix of proteins and ions that serve as the scaffold for calcium phosphate crystals, we recognise as bone. (3)
The temporal and spatial pattern of osteoblast construction of the extracellular scaffold shapes mineral formation, determining the forces a bone can withstand . Here, dynamics at molecular and cellular scales decide outcomes for the organ, and thereby the organism.
Feats of osteoblast engineering continue from early embryonic development until an organism reaches sexual maturity or, in the case of injury, beyond. In the skull, this means the cranial vault can both expand with the underlying brain and heal small fractures. (4)
The capacity to remodel and repair damaged bone has been crucial for adaptation across all vertebrates, particularly in brain development. For example, humans have developed large cortices capable of complex thought partly because the continued growth of the skull vault is not fixed at birth, but is coupled to that of the brain. Diseases affecting bone development significantly impact neurological development, facial structure, and quality of life. (5)
How have we understood skull bone formation?
To treat diseases of the skull (or any other bone) depends on understanding normal processes of development and how these are altered in patients. Developmental biology provides this understanding by studying how tissues develop in embryos. Mouse, zebrafish, and chicken are common systems used to study development, as each shares processes of their skull formation with humans. Until recently, however, tools were limited for tracking dynamic behaviours of bone precursors in whole living tissue, and dynamic information had to be inferred from time series data.
Such studies have shown that the bones of the skull vault first form as small collections of osteoblasts within the layers of the early skull cap. (6) These primordial bones emerge close to the base of the brain and expand as sheets towards the top of the head. How cells behave to expand these bones of the skull vault has remained a mystery.
The cells of the early skull cap that make the bones of the skull vault are particularly enigmatic. These mesenchymal cells, sometimes called fibroblasts, lie deep in tissues. They are mainly flat with spider-like protrusions that extend and retract, transiently interacting with other cells above, below, and on either side, in a matrix of extracellular proteins. It is this dynamism and irregular organisation that have confounded understanding of bone expansion at the level of individual cells.
Cell motion is often ascribed to cell migration
Mesenchymal cells are especially efficient at migrating in directed fashion towards or away from chemical and physical cues. This motion is driven from molecular motors inside each cell, which is analogous to us walking from one place to another. When early experiments suggested that cells in the skull cap moved towards the top of the head (7), it became natural to imagine that early bone cells were doing so through directed collective cellular migration.
Testing this hypothesis began when we discovered a culture method allowing access to deep osteoblasts in the early skull cap for microscopy in living tissue. Finally, we could track fluorescently labelled osteoblasts and ask: How are the bones of the skull vault built? While we found cellular motion consistent with previous work, we discovered another dynamic at play.
Undifferentiated (stem-cell-like) mesenchymal cells that lay in front of differentiated osteoblasts were themselves differentiating, then the cells ahead of those differentiated, and so on, propagating like a wave in the stands of a football stadium. This wave of differentiating osteoblasts spread towards the top of the head, expanding the bone along with it. We sought to understand what generated this differentiation wave and whether it was linked in some way to the cell motion we observed.
The discovery came when we began thinking about the skull mesenchyme and the differentiated osteoblasts as a material generated by the cells collectively. This material contains both cells and the scaffold proteins they secrete. Collagen, the most abundant of bone scaffold proteins, forms networks of fibres that have important material properties depending on density, organisation, and crosslinking. The longer an osteoblast has been differentiated, the more extracellular collagen is locked into a stiff network, forming a stiffness gradient that follows the spatio-temporal wave of differentiation.
Leveraging physics and maths to understand skull formation Mesenchymal cells are particularly responsive to changes in stiffness; in vitro, increasing the stiffness they experience can induce osteoblast differentiation. (8,9) Could differentiation be mechanically coupled to the collagen matrix it produces, forming a mechanical feedback loop?
We drew on mathematical theory, used to describe wave motion in simulations, to test this hypothesis. Not only did our simulations produce a wave of differentiation, but we also found that simulated osteoblasts moved just as we had found in our live imaging experiments. Mechanical feedback between collagen and the differentiation that produces collagen could be sufficient to explain bone expansion.
Our theoretical work left us with a prediction: the slope of the stiffness gradient determines the rate of bone expansion. Through chemical inhibition of an enzyme that promotes stiffening of the collagen network, and embryological manipulations, we confirmed exactly this.
Increasing the stiffness gradient generated bones that were significantly bigger than those of unaltered control embryos. Part of that increased size was because more cells were differentiating at the leading edge of the bone. Differentiation and motion, in contrast, were halted entirely in skull caps where the stiffest region of the bone was removed. (10)
New insights: Spatio-temporal mechanical forces are sufficient to shape tissues
We discovered a new mechanism which can shape tissues. Our data tells us that the mechanical properties of the collagen that differentiated cells secrete establishes a mechanical cue, patterning differentiation in space and time.
This wave is self-propagating because differentiation increases collagen and stiffness locally. Neighbouring undifferentiated cells sense this change and begin producing collagen themselves as they convert to osteoblasts. Thus, the mechanical feedback loop continues. The mechanical byproduct of an expanding collagen gradient is a pressure gradient, capable of pushing cells towards the top of the head. The motion we had ascribed to individual osteoblast migration, could now be explained as cellular motion generated by osteoblasts as a collective.
The ability of a differentiation wave to be regulated by the collective properties of the tissue could also explain how bone expansion scales with the brain in the early embryo. The wave can travel only as long as undifferentiated cells remain to convert into new osteoblasts, thus providing a mechanism for the incredible plasticity of the skull over evolutionary time. Tuning cell numbers and collagen secretion allows skull growth to rapidly adapt to changes in brain size and shape that have defined human evolution.
Future therapies can come from interdisciplinary developmental biology
These insights arise because developmental biology, drawing from all fields of biology, now integrates engineering, physics, and mathematics. We are learning how cells build physical landscapes that instruct fate decisions and movement, and how disease alters these processes. Continued investment in cross-disciplinary collaboration will allow the principles of organ formation to be harnessed to more faithfully reconstruct tissues and improve understanding of bone disease.