
Purple rice, generating genetic diversity, breeding-by-design, large-scale stabilised mutant collection, forward and reverse screening are all discussed here by Prof Dr Apichart Vanavichit from the Rice Science Center, Kasetsart University, Thailand
In this focus on generating renewable genetic variation and enabling next-generation purple rice breeding, we start by looking at the origin of purple rice.
Purple rice has a special place in Asian society, for example, Forbidden rice, is the most delicious rice and only served to Chinese emperors. Most local purple rice varieties are waxy (glutinous) rice, used in special recipes for religious purposes. Scientists believe that purple rice varieties may have first domesticated from the tropical japonica rice group which carried a similar suit of genes controlling anthocyanin biosynthesis. Genetic introgression into local landraces generated two distinct groups of purple rice varieties, small-seed Japonica and large-seed Indica types. As a speciality rice, farmers sporadically cultivated purple rice in several geographically distinct areas in Asia.
Purple rice may become distinct
Purple rice is naturally enriched with antioxidant compounds, vitamins, protein and micronutrients. With limited breeding activities, purple rice germplasm is genetically well preserved and adapts to a specific environment. Several purple rice varieties in Thailand are not highly divergent as judged by the diversity of molecular markers. Also, most purple rice varieties are photoperiod sensitive which limits their growing cycle to the specific season only and are, therefore, susceptible to diseases and insect pests. In modern agriculture, the rapid evolution of diseases and insect pests is the major driving force to lower the productivity of susceptible purple rice. Soon, imminent global warming may force purple rice to become distinct if genetic improvement has not begun.
The quest for new purple rice
Strong interest in breeding purple rice stems from the quest for high nutrient rice. However, breeding effort is constrained by limited access to useful germplasm and genomic information about black rice. In conventional breeding, available germplasm is the most crucial input to enabling breeders to successfully develop new plant varieties. In modern breeding programmes, additional genomics information can speed up the development of high nutritional purple rice and yet it is resistant to biotic and abiotic stresses.
When it comes to rice, the world’s largest germplasm depository is located in the International Rice Genebank at IRRI, where there are more than 132,000 rice accessions and wild relatives from all over the world. Only 5% of the rice germplasm bank at IRRI, however, was utilised for modern rice breeding and only a handful of them were purple rice. There are genetic mechanisms blocking seed setting and recombination in wide hybridization. Incompatibility between genes in the genome and cytoplasmic factors may cause F1-sterility known as wide abortive. As such, crossing and selection with a wild relative have limited successes in self-pollinated species such as rice.
Fasten the rate of spontaneous mutation
While spontaneous mutation slowly generates new favourable alleles for natural selection, induced mutation by radiation and mutagens rapidly alters structural and nucleotide changes to the genome. It is generally believed that induced mutagenesis generates only random alteration on DNA sequences and structural changes at the genome-wide scale. The new paradox of induced mutation was radiation mutagenesis which, not only generates random mutagenesis, but also targets structural genomic changes in more specific events. For example, RD6, the most popular waxy rice from Thailand, is a waxy rice mutant generated by gamma radiation of the iconic Khao Dawk Mali 105, the world-renown Thai Hommali Rice. The sequence variation of its granule-bound starch synthase I, the waxy gene, is similar at the functional sequence variation to any waxy rice.
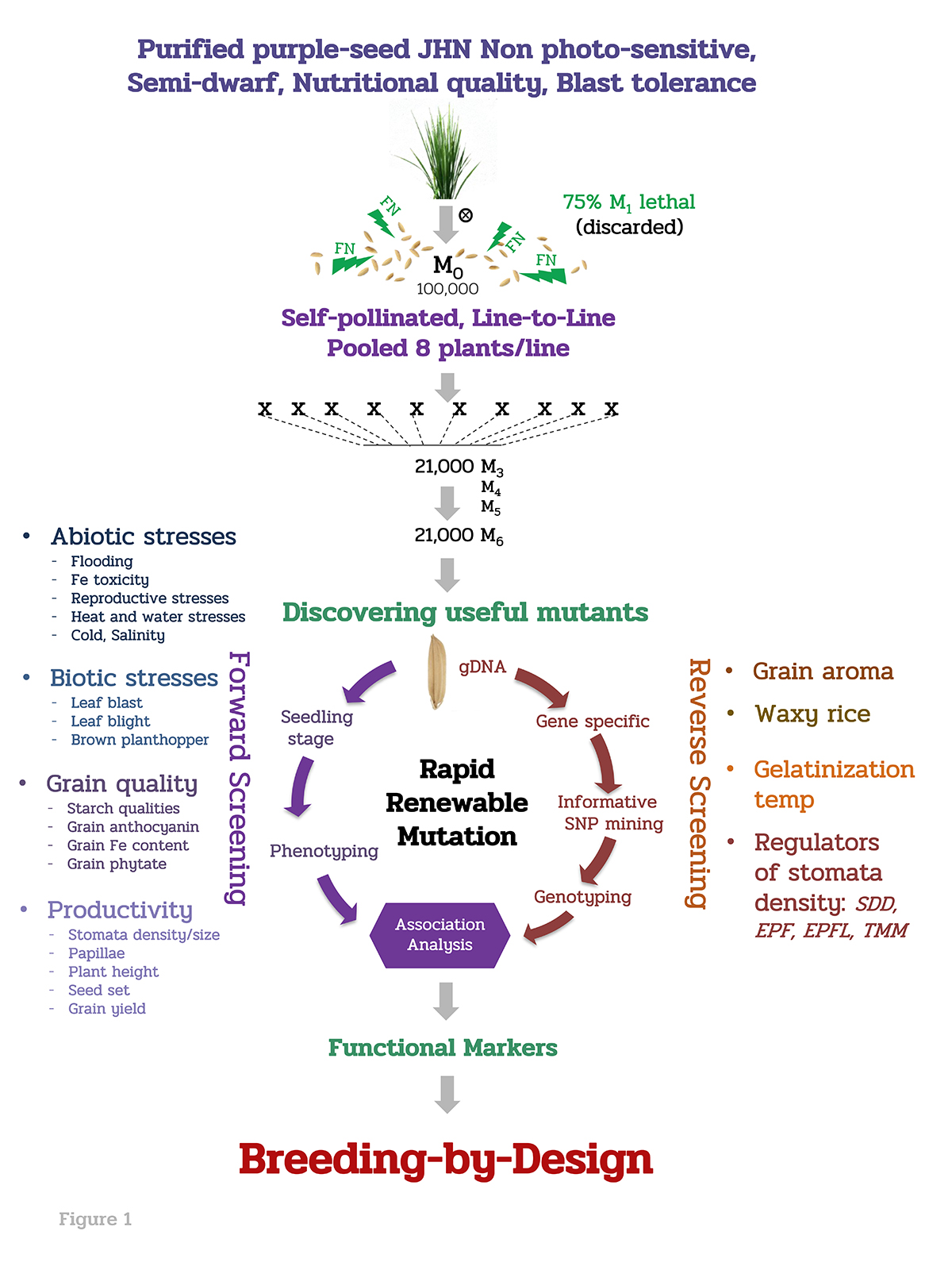
Generating genetic diversity in pigmented rice
To enhance more genetic variability in purple rice germplasm, we treated 33 Gy of fast neutron radiation (FN) on 100,000 genetically pure seeds derived from JHN, a semi-dwarf purple rice with photoperiod insensitivity. The goal is to develop a large purple rice mutant germplasm collection as an alternative genetic resource for breeding climate-ready and high nutrient rice. By non-selective selfing, more than 20,000 M6 stabilized mutant lines were established as a valuable germplasm. Genomic DNA representing each stabilized mutant lines were stocked to facilitate rapid access to specific genomic variation. Collectively, this represents the largest viable, pigmented mutant rice collection for breeding and gene isolation (Figure 1).
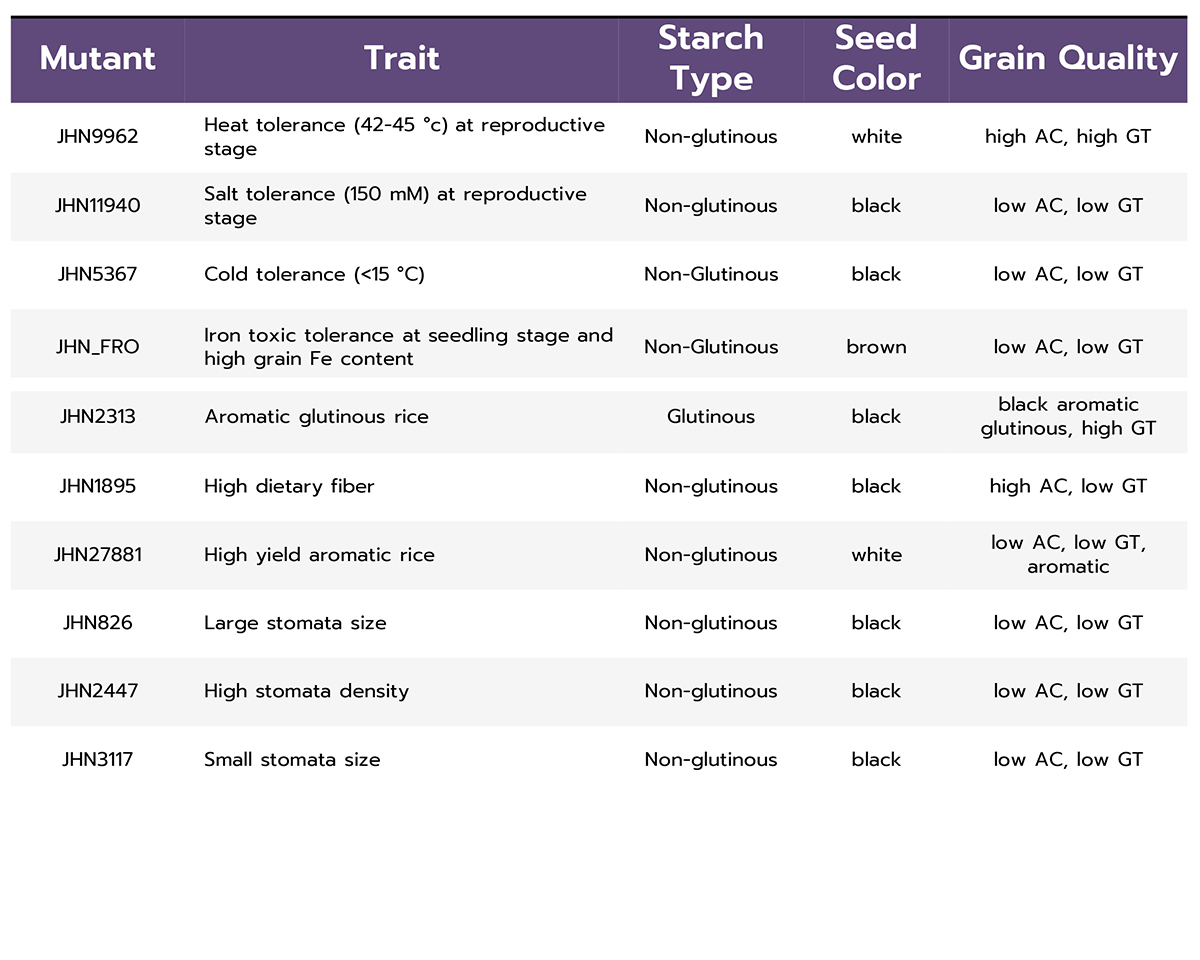
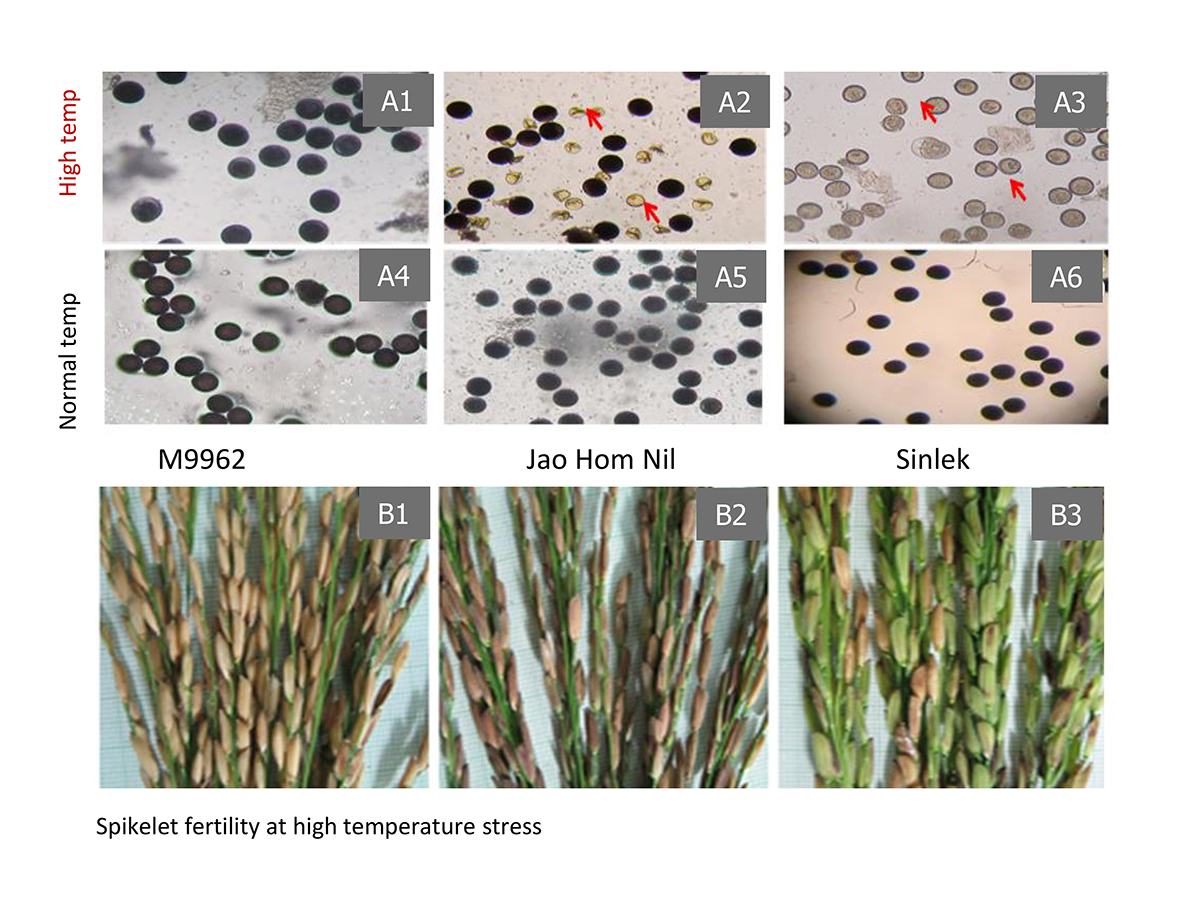
To isolate valuable mutants, large-scale, phenotypic and genotypic screenings (forward and reverse screenings) were carried out, 12 elite mutants were identified for multiple resistance to abiotic, biotic, grain quality and productivity (Table 1). Interestingly, JHN9962 mutant is highly heat-tolerant at the pollination stage, so far (Figure 2). JHN4 mutant is the most submergence tolerance and resistance to brown planthopper carrying mutations on Sub1A and Bph9, respectively (Table 1 and Figure 3).
The genetic variation between individuals is a key phenomenon that underlies natural selection. The factors that drive genetic variation, for instance, by promoting the appearance of spontaneous mutations are not well understood. Over 80% of those were single nucleotide variations (SNV) duplicated with OryzaSNP, the world collection of SNV database. In addition, selection under stresses may generate more specific gain/loss-of-function mutants which carry specific genomic changes and allelic variation similar to what can be found in natural genetic variation. Questions remain concerning which mechanism may induce target genetic variation so rapidly. Consequently, the rapid regeneration of useful genetic diversity empowers breeders to rapidly develop better climate-ready and nutrient-dense rice varieties to cope with the imminent climate changes and specific nutritional needs.

International collaboration
To gain more insight into the new paradigm shift, we are collaborating with Professor Chang-Sheng Wang from the National Chung-Hsing University, Taiwan, to unravel the mechanism of rapid induction of spontaneous mutation by radiation and chemical mutagenesis. His laboratory has successfully developed 4,800 stabilized mutant collections from TNG67 (Japonica) and IR64 (Indica) using NaN3. Valuable mutants were isolated for starch, giant embryo, disease resistance, aromatic grain, and stem brittleness. These valuable mutants will be utilised in breeding-by-design to develop new climate-ready, nutrient-dense rice varieties for the future of sustainable rice production.
*Please note: This is a commercial profile








